 咨询热线:400-065-6886
咨询热线:400-065-6886
咨询热线:400-065-6886
咨询热线:400-065-6886
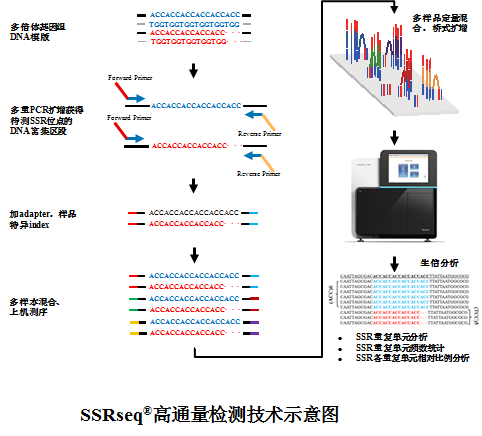
SSRseq®简单重复序列(Simple Sequence Repeat,SSR),也称微卫星(Microsatellite)或短串联重复序列(Short Temdem Repeats,STR),其串联重复的核心序列常为1-6 bp,是一种广泛分布在真核生物基因组中的重复序列。由于其自身具有共显性遗传,多态位点多,信息含量丰富,物种间转移性好,易于检测,重复性好的特点,因此广泛应用于动植物遗传多样性、亲缘关系分析、品种鉴定、分子标记辅助选择、QTL定位等研究。
SSRseq®天昊生物依托自身技术优势,开发出了基于Illumina二代测序平台的SSRseqTM超高通量SSR分型技术。该技术针对目的区域SSR设计多重PCR扩增体系,一个扩增反应体系可以完成16-20个SSR标记的PCR反应,然后对目的片段进行二代测序。此技术可以直接检测到同一SSR位点不同等位基因的序列信息,不但能达到单碱基分辨率,并且能够准确给出每个等位基因SSR重复单元的具体重复次数和不同等位基因的相对比例,不需要人工判读基因型,基于生物信息学分析流程高效、准确分型,适用于二倍体物种,尤其适合异源多倍体物种SSR位点检测。您只需提供样品DNA及SSR位点信息,我们便可帮您完成从实验检测到数据分析整体服务。
| 样本要求 | 位点要求 | 测序策略 | 数据要求 |
|
|
|
|
案例:Genetic diversity analysis of cultivated and wild grapevine (Vitis vinifera L.) accessions around the Mediterranean basin and Central Asia
发表杂志:BMC Plant Biology 影响因子:3.93 发表时间:2018年6月高加索和中国之间的山区被认为是葡萄驯化的中心。尽管中亚在葡萄种植历史上非常重要,但与地中海盆地周围的野生和栽培葡萄相比,关于该地区葡萄遗传变异程度和分布的信息有限。本研究的主要目的是调查高加索、中亚和地中海盆地野生和栽培葡萄种质的遗传多样性和相互关系,了解基因流动、可能的驯化事件和适应性渗入。
利用20个SSR标记对地中海盆地周围和中亚收集的1378种野生和栽培葡萄进行了基因分型。遗传数据经过分析(聚类分析、主坐标分析和群体结构分析)确定了群体分组情况,结果还通过Nei’s遗传距离、成对FST分析和分配检验得到验证。所有这些分析确定了三个遗传群体: G1,来自克罗地亚、法国、意大利和西班牙的野生资源;G2,来自亚美尼亚、阿塞拜疆和格鲁吉亚的野生资源;G3,来自西班牙、法国、意大利、格鲁吉亚、伊朗、巴基斯坦和土库曼斯坦的品种,其中包括来自格鲁吉亚和克罗地亚的少量野生资源。乔治亚的野生资源与同一地区种植的葡萄聚集在一起,但也与西欧聚集在一起,支持乔治亚成为葡萄驯化的古老中心。此外,聚类分析表明,西欧野生葡萄与来自同一地区的栽培葡萄聚在一起,表明栽培的西方proles occidentalis区比来自东欧的野生葡萄对葡萄酒葡萄的早期发展贡献更大。
通过对基因型遗传关系的分析,为地中海盆地和中亚野生和栽培品种间遗传关系提供的新的证据。遗传结构表明有相当多的基因流动,这限制了这两个亚种之间的分化。结果还表明,在野生葡萄被驯化的地区,有着混合祖先葡萄的出现。
; 表1、根据地理来源对1378种栽培和野生资葡萄的分类列表
 --
--; 表2、为1378种不同葡萄基因型多样性指数计算结果
 --
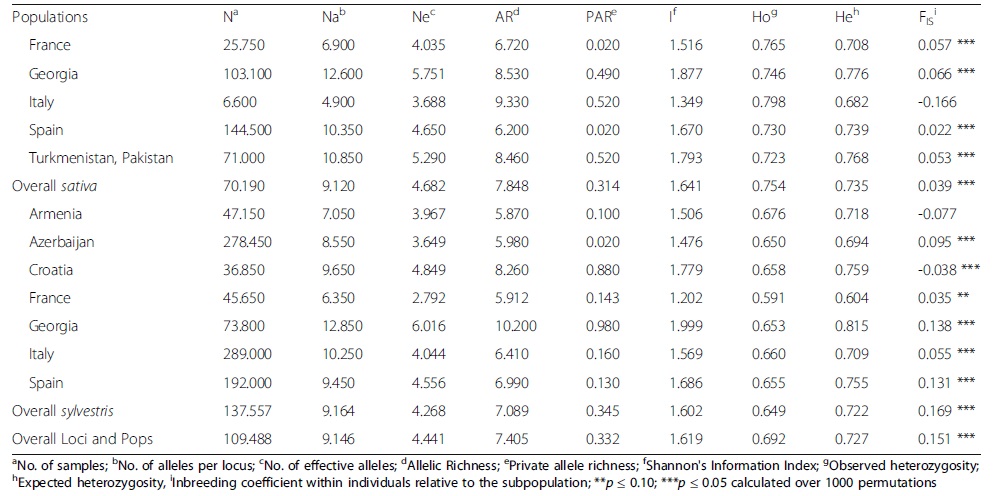
--; 表3、野生和栽培葡萄群体遗传多样性估计
 --
--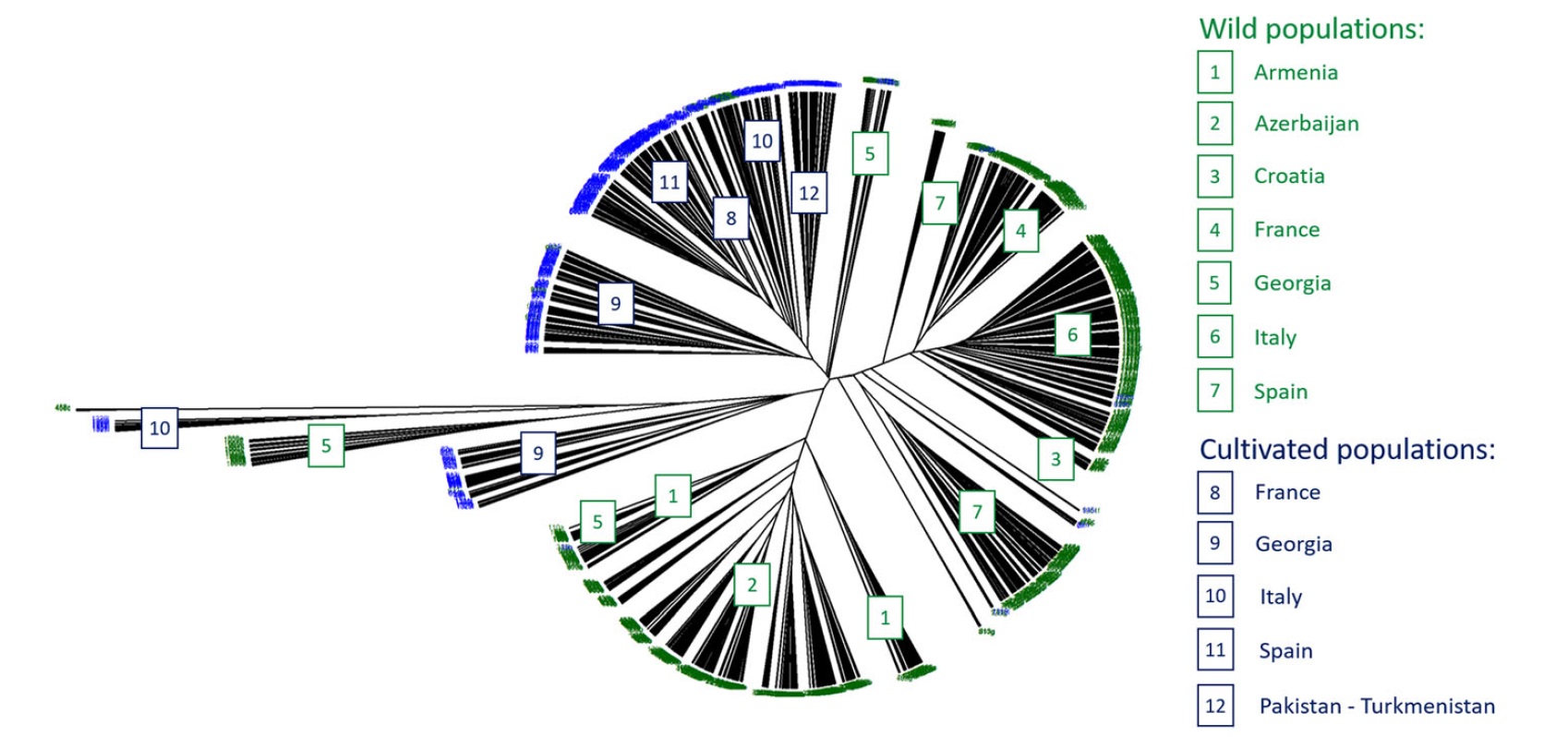
图1、1378种栽培葡萄和野生葡萄NJ聚类图
--

图2、1378种栽培葡萄和野生葡萄PCoA图
--
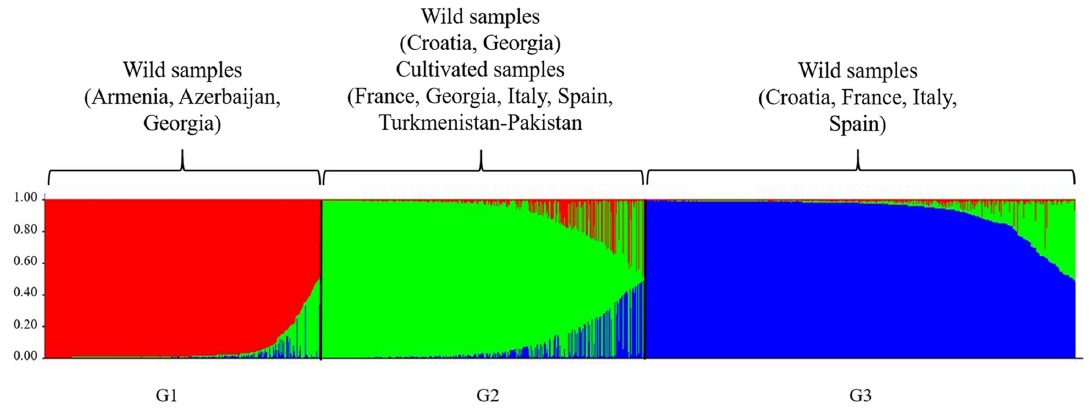
图3、1378种栽培葡萄和野生葡萄群体结构(K=3)分析
--
; 表4、野生和栽培葡萄群体内成对Nei’s遗传距离(对角线以下)和FST值(对角线以上)估计
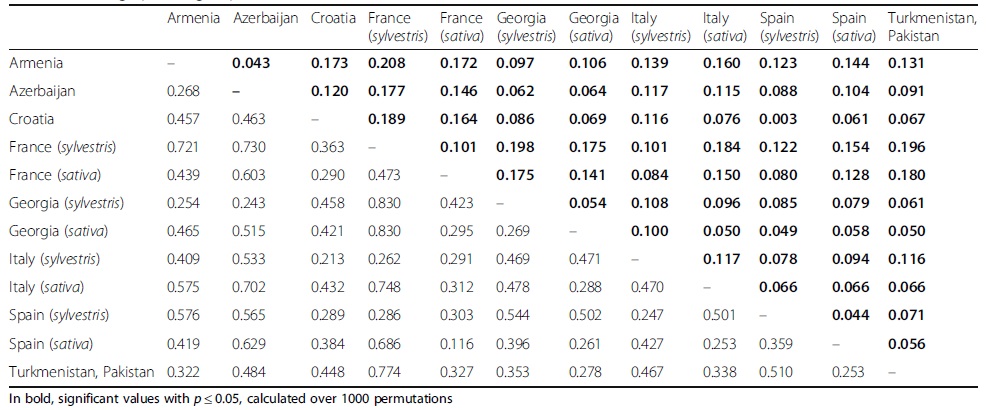
1、本研究对葡萄sativa和sylvestris两个亚种进行了系统分析,SSR基因分型结果表现出高度的多态性和杂合性,说明葡萄亚种内部和亚种之间都存在显著的多样性(表2和3)。
• 来自佐治亚地区的sativa和sylvestris具有最高的有效等位基因数量和等位基因丰富度,这表明该地区是酿酒葡萄多样性的中心。
• 对Ho、He值及FIS的比较结果表明,来自同一野生种群的个体之间存在高度的遗传关系,特别是在西班牙和格鲁吉亚的种群中,这种交配会影响个体和群体的动态,增加近亲交配几率。然而,在随机交配群体中,一些野生群体的FIS值接近于零,这些相反的结果可以用样本大小导致的亚群中等位基因的随机遗传漂移来解释。
• 与其他研究类似,本研究也发现sylvestris样本中观察到的多样性水平的降低。由于森林砍伐和城市化,世界许多地方的sylvestris品种被认为濒临灭绝,支离破碎。人为的和自然的地理障碍也可能导致野生群体在他们的自然栖息地被隔离,并可能导致显著的近亲繁殖,减少不同地理群体内部和之间的基因流动,从而降低杂合度。
• 除了意大利品种外,栽培品种的FIS值接近于零,表明为随机交配。意大利群体的负FIS值表明杂合子过多,但在统计学上没有显著性(表3)。大多数栽培群体中纯合子的缺失表明,它们是由具有不同群体统计学(奠基者效应、瓶颈、扩散)和选择历史的种质组成的。种质收集通常是基因型的混合物。因此,这些集合中的地理群体表现出相对较高的分化水平,导致杂合度高于预期水平。
2、聚类分析和PCoA分析表明,亚种内部和亚种之间的遗传结构和分化具有显著差异(图1和图2)。
• 两种分析都发现西欧野生葡萄和高加索地区野生样品之间有明显的区别,而法国和西班牙野生葡萄关系密切,有着密切的遗传关系。
• 西班牙野生葡萄表现出等级分化,表明相邻种群间的基因流动导致了种群结构的垫脚石模型。或者,等级差异可能是不同地理区域气候差异的结果。
• 克罗地亚的sylvestris与欧洲的sylvestris个体相关,形成了一个基础姐妹群,表明它们有着共同的基因库。包括亚美尼亚、阿塞拜疆和格鲁吉亚在内的外高加索野生葡萄藤组成了一个独特的亚群。
• 在sativa中,观察到来自格鲁吉亚的两组不同的品种,一组是意大利、法国和西班牙品种的姐妹品种(图1),另一组与意大利sativa和格鲁吉亚sylvestris亚组密切相关。这一结果可能表明,中亚和高加索地区的第一批驯化品种在西欧资源中留下了遗传足迹。这种遗传亲缘关系也可能是地中海地区早期育种计划的反映,在那里,具有优良果实的sylvestris或杂交野生藤本植物被用于与驯化品系的杂交。
• 描绘的总体分化模式非常类似于NJ聚类分析。来自格鲁吉亚和亚美尼亚的sylvestris品种中的集群重叠,并与来自格鲁吉亚、巴基斯坦和土库曼斯坦的栽培形式密切相关。
3、STRUCTURE分析支持主要群体之间的差异,而一些群体之间的细微差异,尤其是那些混合群体之间的差异并不明显(图3)。
• 遗传结构的贝叶斯推断表明,这两个亚种之间有相当大的基因流动,并有适度的分化。这些结果表明酿酒葡萄的种植和酿酒促进了野生葡萄的驯化、新品种的创造和葡萄历史早期生长技术的进步。
• 本研究结果表明西欧sylvestris向一些当前西欧品种存在着渗入现象。
• 对高度濒危的栽培葡萄树祖先作为抗病育种基因资源的潜力提供了的支持。鉴于欧亚、北非和亚洲野生葡萄种质和种质在很大程度上还未被开发,它们的鉴定、保存以及生物和非生物抗性和浆果品质的特征对于葡萄酒和葡萄产业的未来非常重要。

